Антагонистические отношения между зрительно-вызванными потенциалами (ЗВП) и гамма силой при синдроме «визуального снега»
Авторы: Себастьян Луна, Даниэль Лай, Элисон Харрис
Принято к публикации 13.10.17
Аннотация статьи
Введение. — Используя «двухимпульсную» адаптационную парадигму, при которой два стимула представлены в быстрой последовательности, в этом исследовании рассматриваются нейронные механизмы, лежащие в основе потенцирования зрительно-вызванного потенциала (ЗВП) при синдроме «визуального снега».
Справочная информация. — Синдром «визуального снега» – это постоянное нарушение восприятия, характеризующееся быстрыми мерцающими точками во всем поле зрения. Так как с этим синдром связано состояние мигрени с аурой, было высказано предположение, что «визуальный снег» возникает из-за аномальной чувствительности к нейронам, о чем свидетельствует отсутствие типичного «привыкания» зрительно-вызванного потенциала к повторной визуальной стимуляции. Однако нейронные механизмы, которые лежат в основе этого эффекта, остаются невыясненными. Предыдущие эксперименты с «двойным импульсом» предполагают, что типичное привыкание «ЗВП» отражает разрушительные нейронные колебания гамма-диапазона (50-70 Гц) , которые, возможно, обусловлены тормозящими вставочными нейронами. Учитывая, что мигрень ассоциируется с уменьшенным ингибированием коры головного мозга, мы предполагаем, что «визуальный снег» может также отражать уменьшенную ингибирующую активность, которая приводит к уменьшению мощности гамма-излучения после первоначальной визуальной стимуляции и сопутствующего потенцирования последующего ответа ЗВП.
Методы. — Мы сравнили ответы ЗВП на двух импульсную адаптацию у 22-летнего мужчины с двухлетней историей «визуального снега» по сравнению с контрольной группой по возрасту и полу (N (количество) = 5). У пациента не имелось сопутствующего диагноза эпизодической мигрени или мигрени с аурой, и у контрольной группы не было личного или семейного (наследственного) анамнеза мигрени.
Результаты. — В отличие от характера привыкания, наблюдаемого в контрольной группе, «визуальный снег» был связан с постоянным потенцированием ответа ЗВП. В соответствии с нашими прогнозами, частотно-временный анализ показал снижение мощности гамма-диапазона после первоначального стимула при «визуальном снеге» относительно контрольной группы.
Выводы. — Эти результаты подтверждают антагонистическое взаимодействие между гамма-мощностью и быстрой нейронной адаптацией, проливая новый свет на нейронные механизмы потенцирования ЗВП при синдроме «визуального снега».
Введение
Положительные визуальные феномены, при которых пациенты испытывают иллюзии или искажения зрительного восприятия, являются результатом ненормальной нейронной реакции при различных патологических состояниях. Несмотря на то, что нейронные механизмы положительных визуальных феноменов широко изучались в контексте мигрени с аурой, мало известно о лежащей в основе патологии визуальном расстройстве при синдроме «визуального снега. Люди с «визуальным снегом» сообщают о стойком и постоянном визуальном нарушении во всем поле зрения, которое характеризуется быстрым мерцанием мелких точек, напоминающих эффект «видеошума» или «зернистость» изображения».
Как и мигрень, «визуальный снег» предположительно возникает из-за изменений чувствительности нейронов к сенсорной стимуляции. Измерения зрительно-вызванных потенциалов (ЗВП) у здоровых людей обычно показывают физиологическое привыкание и уменьшение нейронного ответа при повторных стимуляциях.И, напротив, у пациентов с мигренью, зрительно-вызванные потенциалы при повторной стимуляции часто потенцируются или увеличиваются. В соответствии с этой идеей недавнее исследование сообщило о потенцировании зрительно-вызванных потенциалов при синдроме «визуального снега». Однако эти результаты были поставлены под сомнение в связи с наличием мигрени с аурой в анамнезе,а также в связи с более общими опасениями по поводу достоверности привыкания зрительно-вызванных потенциалов при мигрени.
Здесь мы тестировали привыкание зрительно-вызванных потенциалов у 22-летнего мужчины, страдающего «визуальным снегом» без сопутствующей мигрени, что сделано с целью исключения мигрени с аурой в качестве негативного фактора. Кроме того, мы расширяем предыдущие исследования путем измерения привыкания ЗВП с использованием быстрой парадигмы адаптации “с двойными импульсами”, в которой два стимула (S1 и S2) представлены с переменной асинхронностью наступление стимула (SOA (АНС)). Адаптация к двухимпульсным «представлениям» наблюдалась для сигналов, возникающих как из начала зрительной зоны коры головного мозга, так и для зон восприятия более высокого порядка, и может быть представлена в виде парных презентаций разных изображений, предполагая тот факт, что адаптация не зависит только лишь от физических свойств стимула низкого уровня. Вместо того, чтобы индексировать прямое снижение (уменьшение) нейронной активности, этот адаптационный эффект, по-видимому, отражает нарушение S2-реакции синхронизированными колебаниями гамма-диапазона (50-70 Гц), которые предшествуют началу стимула S2, причём с большим подавлением вызванного S2-ответа, связанного с более высокой гамма-мощностью pre-S2 (трудно интерпретировать pre-S2 в переводе).
Один из возможных нейронных механизмов разрушающего действия пре-S2-гаммы связан с предыдущей работой, в которой показано, что синхронные кортикальные гамма-колебания зависят от активности тормозных интернейронов (вставочных нейронов). Интересно, что ингибирование зрительной зоны коры головного мозга, по-видимому, уменьшается при мигрени, о чем свидетельствуют исследования с использованием метаконтраста (метаконтрастной маскировки) и неинвазивной стимуляции головного мозга. Таким образом, мы выдвигаем гипотезу о том, что при «визуальном снеге» снижение ингибирования зрительной зоны коры головного мозга приведет к уменьшению синхронных гамма-колебаний (т. е. меньшей мощности), предшествующих стимулу S2, относительно нормальных уровней, с сопутствующим отсутствием привыкания ЗВП.
Методы
Пациент – мужчина-правша 22 лет с 2-ух летней историей (длительность заболевания) синдрома «визуального снега». У него отмечаются некоторые из недавно предложенных диагностических критериев «визуального снега», главным из которых является наличие постоянно мелькающих мелких точек по всему полю зрения (в этом предложении имеется ввиду, что он подходит под эти критерии).
Этот симптом сохраняется как при наличии света, так и после закрытия глаз. Пациент также испытывает несколько связанных положительных зрительных феноменов, включая палинопсию, никталопию, фотопсию, фосфены, энтопический феномен синего поля (для справки - феномен Ширера) и шум в ушах. Все симптомы развивались в течение двух недель и с тех пор сохраняются без признаков ремиссии. У пациента нормальная острота зрения и строение глаз, при неврологических и нейровизуальных исследованиях не было обнаружено никаких аномалий.
Учитывая, что наличие пяти приступов головной боли мигренозного типа в прошлом являются достаточными для диагностики эпизодической мигрени, важно оценивать не только количество приступов, но и степень их проявления. Пациент испытал одну атаку мигрирующего типа с типичными симптомами ауры мигрени, которая произошла за 6 лет до проведения данного исследования. Других приступов мигрени пациент не испытывал, поэтому не может претендовать на установление диагноза эпизодической мигрени. Тем не менее, существует семейная (наследственная предрасположенность) история мигрени с аурой на материнской стороне.
Семь гендерных и возрастных контролей (возраст 20-24 лет, средний возраст = 21,1) были набраны из сообщества колледжей, из которых двое были исключены из-за проблем с записью ЭЭГ. У всех участников контроля было нормальное или исправленное зрение, а также личная или семейная история мигрени. Информированное согласие было получено от всех участников, и исследование было одобрено экспертным советом колледжа.
В «двухимпульсном» эксперименте по адаптации два стимула были представлены по 17 мс каждый с четырьмя асинхронными наступлениями стимула: 50, 67, 117 и 217 мс (рисунок 1А). Стимулы состояли из 50 высококонтрастных черно-белых линий (4,6 ° × 4,6 ° зрительного угла), представленных на сером фоне с центральной точкой фиксации (рисунок 1B). Испытуемые наблюдали за редким целевым изображением шахматной доски (10% от общего количества испытаний), появление которой случайным образом смешивалось с экспериментальными испытаниями. Все испытания были запрограммированы и отображены в Matlab (Mathworks, Natick, MA, USA) с использованием программного обеспечения для презентации стимулов PsychToolbox.
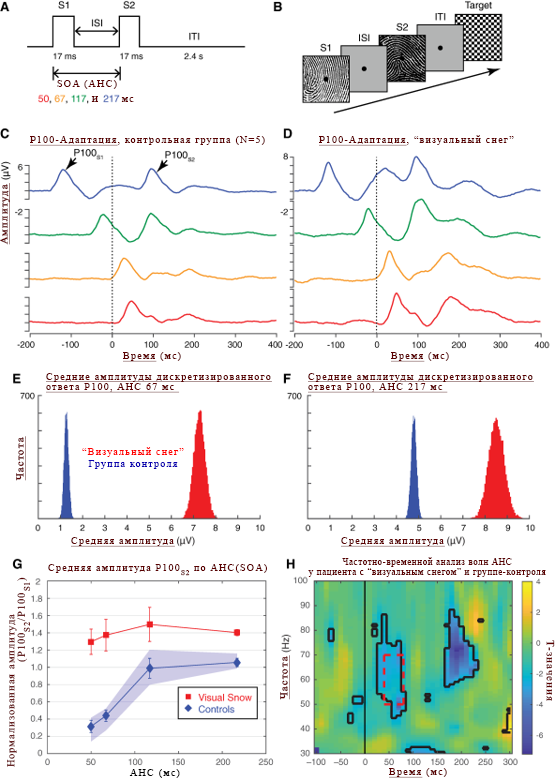

Пояснения к рисунку 1.
(А) Схема парадигмы адаптации с «двойным импульсом». Два стимула, S1 и S2, представлены в течение 17 мс каждый с переменным интерстимулярным интервалом (ИСИ)* от 33 до 200 мс. Асинхронность наступления стимула (SOA (АНС)) – 50, 67, 117 и 217 мс.
*с переменным интервалом между стимулами – можно интерпретировать ещё так.
(B) Двухимпульсная адаптация раннего визуального компонента P100. На каждом испытании 2 из 50 высококонтрастных черно-белых (отпечатки пальцев) стимулов были выбраны и представлены парами, причем каждый образец служил как изображение S2 – дважды за состояние (100 испытаний на состояние). Отличающимся (неидентичным) изображением было случайно выбрано S1. Участникам необходимо было ответить на появление редкого целевого изображения (в виде шахматной доски), которое было использовано в 10%-х испытаний.
(C-D) Р100-адаптация в контрольной группе и у пациента с синдромом «визуального снега» через четыре асинхронных наступления стимула (цвета выделены на изображении А**). Наиболее усреднённые формы волн фиксируются до начала стимула S2 (пунктирная линия, 0 мс), а ответ Р100S2 начинается примерно через 100 мс после начала стимула. Следует обратить внимание, что при более короткой асинхронности наступления стимула, P100S1 визуализируется в момент, предшествующий P100S2. Наиболее усреднённые значения амплитуды волн у пациента с «визуальным снегом» изображены на рисунке (D).
(E-F) Средние амплитуды повторного дискретизированного ответа P100S2 для условий АНС (E) 67 мс и (F) 217 мс. Средние амплитуды вычислялись для каждого испытания у каждого субъекта в течение 20 мс с учётом средней пиковой задержки, а распределения загрузочных средств были получены из 1000 выборок с заменой пробных данных внутри объекта.
(G) Среднее значение амплитуды P100S2 согласно АНС у пациента с «визуальным снегом» (красные квадраты) по сравнению с контрольной группой (синие ромбы). Значение погрешности представляет стандартную ошибку среднего. Заштрихованная синяя область указывает на 95% доверительных интервалов, определяемых путем BootsTrap (10 000 перестановок(переменных)).
(H) Частотно-временной анализ коротких и длинных волн асинхронности наступления стимула у пациента с синдромом «визуального снега» и контрольной группе. Красная пунктирная рамка: представляющее интерес частотно-временная область, в которой показаны различия гамма-мощности. Черные контуры показывают области, в которых значения t были значительными; исправлены для множественных сравнений с использованием ложной скорости обнаружения (FDR(ЛСО)).
**имеется ввиду цвета времени асинхронного наступления стимула (50 мс – красный – он же на рисунке С выделен красным.)
Данные ЭЭГ были собраны с использованием 128-канальной системы BioSemi ActiveTwo (Biosemi B.V., Амстердам, Голландия). Оффлайн-обработка данных выполнялась с использованием инструментальной панели EEGLAB (ЭЭГЛАБ),включая повторную выборку до 500 Гц, повторную привязку к среднему значению, линейное детрендирование, фильтрацию высоких частот на 1 Гц и выбор временных промежутков, привязанных к стимулу S1 (от -500 мс до 800 мс). Искусственный шум был идентифицирован и удален с помощью независимого анализа компонентов с помощью слепой идентификации второго порядка и промежуток времени в 600 мс, привязанных к стимулу S2 (от -200 мс до 400 мс). Предварительная обработка для временной частоты выполнялась по этой же аналогии, но исключая повторную привязку и фильтрацию данных.
Для измерения адаптации зрительно-вызванных потенциалов (ЗВП) мы изучили Р100-ответ, наступающий примерно через 100 мс после стимуляции на задних датчиках. Для каждого участника индивидуальные датчики «заинтересованности»(здесь проблемно перевести (ИДЗ)) (СОИ) были определены через 90-110 мс после стимула на задних датчиках c учётом амплитуды ЗВП S1-стимула, с использованием порога амплитуды z, набравшей ≥ 1,5. Затем определялась локальная амплитуда пика и латентность для ответа S2 для каждого участника с использованием 10-точечной (20 мс) области в инструментальной панели ERPLAB для Matlab. Для обеспечения того, чтобы на эти средние значения уровня субъекта не было непропорционального влияния малого количества отдаленных значений, мы эмпирически оценили среднюю амплитуду ответа P100S2 для области в 20 мс (10 мс до и 10 мс после) с учётом средней пиковой задержки в условиях асинхронного наступления стимула (67 мс и 217 мс) на основе отдельных пробных данных с использованием процедуры BootsTrap (1000 выборок). Оценки BootsTrap вычислялись отдельно через элементы управления, чтобы сохранить изменчивость между субъектами. Степень адаптации была количественно определена путем отношения ответа P100 на стимул S2 к амплитуде предыдущего ответа S1 (P100S2 / P100S1). Доверительные интервалы BootsTrap для нормализованного ответа P100S2 были построены с использованием 10,000 переменных (перестановок), пересчитывающих исходные данные субъекта с заменой.
Частотно-временной анализ проводился с помощью набора инструментов FieldTrip для Matlab с использованием вэйвлета Morlet (ширина = 7). Частотно-временной анализ проводился индивидуально для каждого участника на все испытания, усредняя по индивидуально определенным СОИ(ИДЗ) участника. Сначала мы вычисляли разницу в мощности между коротким и длинным СОИ, определяемой 67 мс против 217 мс соответственно. Затем мы сравнили разницу коротковолнового ИДЗ у пациента с синдромом «визуального снега» по сравнению с контрольной группой участников при помощи двухстороннего t-теста независимых образцов с непараметрической статистикой переменных Монте-Карло (10000 повторений). Результаты были скорректированы для множественных сравнений с помощью ложной скорости обнаружения (ЛСО, (FDR). Основываясь на наших независимых гипотезах, мы предположили, что значительное снижение гамма-мощности будет видно в временном промежутке, предшествующем ответу P100S2 (пост-симул от 40 до 80 мс) в диапазоне частот от 50 до 70 Гц.
Результаты
Наиболее усреднённые формы сигналов в контрольной группе (рис. 1С) и у пациента с «визуальным снегом» (рис. 1D) выявлены узнаваемые компоненты Р100 в ответ на стимулы S1 и S2. Характер привыкания в контрольной группе соответствовал ранее полученным результатам об уменьшении адаптации при более длинных АНС (SOA) с полным восстановлением амплитуды P100 в среднем на 117 мс SOA (АНС) (рис. 1C). Напротив, через два отдельных сеанса записи зрительный снежный пациент показал характерный характер адаптации, характеризующийся заметным потенцированием ответа P100S2 (рис. 1D). В отличие от этого, на двух отдельных сеансах, пациент с «визуальным снегом» показал отличительную картину адаптации, характеризующуюся выраженным потенцированием реакции P100S2 (рис. 1D). Хотя измерение реакции P100S2 на более коротких АНС (SOA) осложнялось атипичным появлением формы ЗВП, мы сосредоточились на первом положительном пике после P100S1 (средняя латентность: 50 мс SOA (АНС) = 181,7 мс; 67 мс SOA (АНС) = 172 мс) из-за сообщения пациента о длительном остаточном изображении после визуальной стимуляции, а также наблюдения, что даже для SOA(АНС) в 117 мс средний ответ-P100S2 пациента с «визуальным снегом» показал увеличение пиковой задержки по сравнению с задержками в группе контроля (t (4) = -8,47, P = 0,001, один образец t-теста).
Чтобы гарантировать, что различия между средними формами сигналов в контрольной группе и у пациента с «визуальным снегом» не были обусловлены малым количеством отдаленных испытаний, мы дополнительно реконструировали распределение средних амплитуд P100S2 для двух из условий SOA(АНС), 67 мс (рис. 1E) и 217 мс (рис. 1F), используя процедуру повторной выборки BootsTrap для отдельных пробных данных (1000 выборок).
Сравнение среднего значения амплитуды P100, вызванной повторной выборкой, сравнивая контрольную группу (синее обозначение) и пациента с «визуальным снегом» (красное обозначение), показало, что два распределения были полностью разделены, причем более высокие средние амплитуды для пациента с «визуальным снегом») были независимо от состояния SOA (АНС).
Рассматривая весь спектр тестируемых SOA (АНС), эффекты адаптации у пациента с «визуальным снегом» были резко уменьшены по сравнению с контрольной группой (рис. 1G). Bootstrap пересчитал 95% доверительных интервалов (рис. 1G, затемненная синяя область) и не выявил перекрытия между контрольной группой и пациентами с «визуальным снегом». Чтобы подтвердить этот результат статистически, мы провели дисперсионный анализ у пациента с «визуальным снегом» в качестве фактора между субъектами и SOA(АНС) в качестве повторных измерений. Степени свободы (?-устоявшееся определение) были скорректированы с целью исправления нарушений сферичности по среднему оценочному эпсилону от Гринхауза-Гейссера и Гюни-Фельдта. Анализ выявил существенный основной эффект SOA(АНС) (F (2.05,8.2) = 8,79, P = 0,009) у субъектов, а также значительное взаимодействие SOA(АНС) x «визуальный снег» (F (2,05,8,2) = 4,33, P = 0,05 ) и эффект у пациента с визуальным снегом: (F (1,4) = 23,2, P = 0,009).: (F (1,4) = 23,2, P = 0,009).
Мы также предположили, что отсутствие привыкания будет коррелировать с уменьшенной гамма-мощностью во времени, которое предшествовало второму стимулу. Для каждого участника мы вычислили гамма-мощность, связанную с длинным и коротким SOA(АНС) (217 против 67 мс) от -100 до 400 мс после начала второго стимула. Мы предположили, что в момент, непосредственно предшествующий началу реакции S2, в контрольной группе будет наблюдаться положительный эффект (т. е. большая разрушительная гамма-мощность в коротком состоянии SOA(АНС)), в то время как у пациента с «визуальным снегом» будет отрицательный или нулевой эффект. Поэтому, сравнивая пациента с «визуальным снегом» и контрольную группу, мы ожидали отрицательный эффект в пределах 50 и 70 Гц, примерно через 40-80 мс после начала стимуляции S2 (рис. 1F, красная пунктирная область).
В соответствии с этим прогнозом, независимый T-тест с использованием непараметрической статистики переменных Монте-Карло (10 000 переменных) обнаружил значительные отрицательные значения t, перекрывающиеся с приоритетным частотно-временным промежутком (окном) (рис. 1F).
Обсуждения
Несмотря на растущее осознание синдрома «визуального снега», как отдельного от мигрени с аурой расстройства, нейронные основы этого состояния остаются неясными. Здесь мы рассмотрели этот вопрос, измерив привыкание к ответу зрительно-вызванных потенциалов с использованием парадигмы «двойного импульса». Наши результаты соответствуют предыдущим результатам потенцирования ЗВП при синдроме «визуального снега», продолжая демонстрировать этот эффект при отсутствии сопутствующей мигрени с аурой. Этот эффект потенцирования наблюдался во всем диапазоне испытанных асинхронных стимуляций и сохранялся у участников контрольной группы. Поскольку все условия SOA были случайно «переплетены», а не представлены в последовательных блоках, наблюдаемая разница вряд ли будет объяснена негативными факторами, такими как нарушение внимания или повышенная усталость.
Более того, наши данные показывают новую механистическую связь между потенцированием VEP(ЗВП) и нервными колебаниями в гамма-диапазоне. В соответствии с предыдущими исследованиями мы наблюдали антагонистическую зависимость между гамма-мощность до S2 и последующей вызванной реакцией с уменьшенной гамма-синхронизацией в группе пациента с «визуальным снегом». Хотя происхождение этого эффекта еще не полностью понятно, тормозные вставочные нейроны (интернейроны), по-видимому, играют ключевую роль в «работе» гамма-колебаний, тем самым обеспечивая кортикальные сенсорные реакции. В свете этого, сообщалось о гиперметаболизме в зрительной зоне коры головного мозга у пациентов с «визуальным снегом», что согласуется со сниженными тормозящими процессами.
Наши результаты сходятся с литературными данными по поводу «привыкания» к зрительно-вызванным потенциалам при мигрени, которая, по данным разных авторов, не только приводит к снижению ингибирования, но и увеличению возбудимости коры или снижению уровня пре-активации. Несмотря на то, что мы не можем исключить альтернативные объяснения потенцирования VEP, наблюдаемые здесь, наши результаты подчеркивают важную роль для временно-частотного анализа при интерпретации вызванных потенциальных данных. В частности, появление антагонистической гамма-синхронизации во временном окне после ответа P100S1 указывает на специфическую роль локальных рекуррентных цепей в опосредованной адаптации с «двойным импульсом». Низкая синхронность нейронных колебаний в этот период времени может взаимодействовать с со сниженным исходным уровнем активности и/или нейрональной гипервозбудимости, чтобы обеспечить большую синхронизацию ответа на последующий стимул S2, что в результате приводит к наблюдаемому здесь потенцированию VEP.
В конце хотелось бы отметить, что, хотя наши данные в значительной степени согласуются с весомой научной литературой по «двухимпульсной» адаптации, для подтверждения этих результатов потребуется проведение дальнейших исследований. Например, несмотря на то, что в настоящее время у пациента с «визуальным снегом» отсутствуют симптомы мигрени с аурой, в прошлом имел приступ мигрени с аурой и, следовательно, такой приступ может повторить в будущем. В более общем плане, сравнение большего количества пациентов с контрольными мигренями и аурой, а также оценка других зрительно-вызванных потенциалов будет необходимо для устранения неврологических факторов, способствующих развитию «визуального снега».
Выводы
В совокупности эти данные дают новое представление о физиологической основе позитивных визуальных феноменов у пациентов с синдромом «визуального снега». Будущие исследования, изучающие адаптацию с «двойным импульсом» с привлечением большего количества пациентов и дополнительных визуально вызванных компонентов, могут попытаться пролить свет на физиологические соотношения при этом изнурительном состоянии.
Благодарности
Авторы благодарят Алену Янг за ее проницательные комментарии к предыдущей версии рукописи.
Авторское заявление
Категория 1
(a) Концепция и дизайн
Себастьян Луна, Элисон Харрис
(b) Поиск материала (данных)
Себастьян Луна, Даниэль Лай, Элисон Харрис
(c) Анализ и интерпретация данных
Даниэль Лай, Элисон Харрис
Категория 2
(a) Составление рукописи
Себастьян Луна, Даниэль Лай, Элисон Харрис
(b) Пересмотр его интеллектуального содержания
Себастьян Луна, Элисон Харрис
Категория 3
(a) Окончательное утверждение завершённой рукописи
Себастьян Луна, Даниэль Лай, Элисон Харрис
***По всему тексту слово "стимулы" можно интерпретировать как раздражители.